


Anti-Lin28: Mouse Lin28 Antibody |
 |
BACKGROUND Lin28 is an evolutionarily conserved RNA-binding protein and is widely expressed in early-stage embryos, with expression decreasing and becoming restricted to a limited number of tissues as embryonic development proceeds. The skeletal and cardiac muscles are among the few adult tissues in which Lin28 expression persists. In human and mouse embryonic stem (ES) cells, Lin28 is abundantly expressed, and expression is down-regulated upon induction of differentiation. The functional significance of Lin28 is further highlighted by its ability to enhance the efficiency in reprogramming human fibroblasts to pluripotency. Lin28 is among four factors (including Oct4, Sox2 and Nanog) that together reprogram human fibroblasts to pluripotency.
Predominantly cytoplasmic, Lin28 can be found in a variety of complexes including messenger ribonucleoprotein particles (mRNPs), polysomes, P-bodies, and stress granules. Functionally, Lin28 associates with RNPs containing IGF-2 mRNA and promotes its translation in differentiating muscle cells. Moreover, Lin28 acts to modulate mouse ES cell growth likely in part by effecting the expression of cell cycle genes (including cyclins A and B and cdk4) at the translational level.1 It also inhibits the processing of let-7 microRNAs that have been implicated in the regulation of cell growth and differentiation in embryonic cells. This inhibition requires Lin28 specifically to bind the terminal loop region of let-7 precursors and recruits ZCCHC11/TUT4 uridylyltransferase, leading to the terminal uridylation of pre-let-7. Uridylated pre-let-7 miRNAs fail to be processed by Dicer and undergo degradation.2 In addition, it was shown recently that Lin28 can interacts with RNA helicase A (RHA) to regulate Oct4 expression at post-transcriptional level.3 It also promote the expression of the H2a gene (and perhaps also other replication-dependent histone genes) at the posttranscriptional level in order to coordinate histone production with the unique proliferative properties of embryonic stem cells. Thus, Lin28 can specifically regulate gene expression through multiple mechanisms.
Expression and function of Lin28 itself was regulated by various signaling pathways. It was shown that Src mediated cell growth, adhesion, and communication through regulating the expression of Lin28.4 Raf kinase inhibitory protein (RKIP) negatively regulates the MAP kinase (MAPK), G protein-coupled receptor kinase-2, and NF-κB signaling cascades. RKIP has been implicated as a metastasis suppressor for prostate cancer. RKIP represses invasion, intravasation and bone metastasis of breast tumor cells in part through a signaling cascade involving inhibition of MAPK, Myc, and LIN28, leading to induction of the microRNA let-7 and downregulation of its targets.5 In addition, LIN28 was overexpressed in primary human tumors with poor clinical prognosis. The overexpression is linked to repression of let-7 family miRNAs and derepression of let-7 targets.6
REFERENCES
1. Xu, B. et al: RNA 15:357-61, 2009
2. Piskounova, E. et al: J. Biol. Chem. 283:21310-14, 2008
3. Qiu, C. et al: Nucleic Acid Res. 38:1240-8, 2010
4. Khusial, P. R.et al: Cell Communication & Adhesion 15: 407 – 409, 20085.
5. Dangi-Garimella, S. et al: EMBO J. 28:347-58, 2009
6. Viswanathan, S.R. et al: Nature Genet. 41:843-8, 2009
2. Piskounova, E. et al: J. Biol. Chem. 283:21310-14, 2008
3. Qiu, C. et al: Nucleic Acid Res. 38:1240-8, 2010
4. Khusial, P. R.et al: Cell Communication & Adhesion 15: 407 – 409, 20085.
5. Dangi-Garimella, S. et al: EMBO J. 28:347-58, 2009
6. Viswanathan, S.R. et al: Nature Genet. 41:843-8, 2009
Products are for research use only. They are not intended for human, animal, or diagnostic applications.
Параметры
Cat.No.: | CP10153 |
Antigen: | Purified recombinant human Lin28 fragments expressed in E. coli. |
Isotype: | Mouse IgG1 |
Species & predicted species cross- reactivity ( ): | Human, Mouse, Rat |
Applications & Suggested starting dilutions:* | WB 1:1000 IP n/d IHC 1:200 ICC 1:200 FACS n/d |
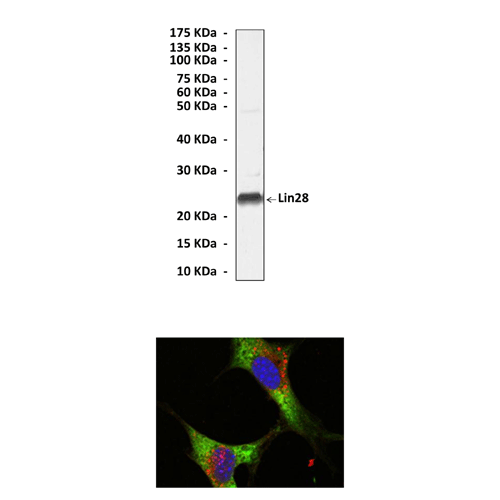
Predicted Molecular Weight of protein: | 23 kDa |
Specificity/Sensitivity: | Detects endogenous Lin28 proteins without cross-reactivity with other family members. |
Storage: | Store at -20°C, 4°C for frequent use. Avoid repeated freeze-thaw cycles. |
*Optimal working dilutions must be determined by end user.
Документы

Информация представлена исключительно в ознакомительных целях и ни при каких условиях не является публичной офертой



