


Anti-WIF1: Mouse WIF1 Antibody |
 |
BACKGROUND The Wingless-type (Wnt) family consists of a group of signaling molecules that is extensively involved in developmental processes and oncogenesis. The proto-oncogenic effects of Wnt genes were discovered, and numerous reports have demonstrated aberrant activation of the Wnt signaling pathway in disparate human cancers. In addition to Wnt ligands and receptors, three classes of secreted antagonists of the Wnt pathway have been identified: secreted Frizzled-related protein (sFRP) family, Dickkopf (Dkk) family, and Wnt inhibitory factor 1 (WIF1). These antagonists can modulate Wnt signaling either by binding to Wnt ligands or by binding to the LRP5/6 co-receptor, leading to receptor endocytosis. WIF1 binds to Wnt proteins and inhibits their activity. Recently, the WIF1 gene was shown to be downregulated in various humanr tumors. It was shown that decreased WIF1 expression correlated with methylation of CpG dinucleotides within the WIF1 promoter. Numerous tumor suppressor or growth regulatory genes have been shown to undergo aberrant de novo methylation and transcriptional silencing in human tumors. Site-specific DNA methylation is often an early event, as demonstrated in tumor types with a well-defined pattern of progression, such as colon cancer, and is now widely regarded as one of the ‘hits’ in the Knudsen hypothesis leading to tumor suppressor gene inactivation.1
The commonly described effect of WIF1 on cancer cells is the inhibition of cancer cell growth. The Wnt/β-catenin pathway has been shown to determine the proliferation/differentiation balance through its regulation of G1-S transition in several cellular systems. The G1-S transition in cell cycle is driven mainly by cyclin-dependent kinase (CDK) 2 that is controlled by abundance of CDK inhibitors: p21/WAF1 (p21) and p27/Kip1 (p27). The regulation of G1-S transition is physiologically required for cell fate determination—a cell undergoing apoptosis, proliferation, or differentiation. However, during oncogenic transformation, G1-S transition is deregulated by enhanced oncogenic growth signaling and/or by loss of tumor suppressors, which then leads to overgrowth of transformed cells. Some Wnts (e.g., Wnt1) have shown oncogenic activities in both mouse models and cell cultures. The activation of the Wnt/β-catenin pathway by Wnts elicits specific target genes (e.g., c-myc and cyclin D1) for cell cycle regulation and growth. It is possible that the secreted Wnt antagonist WIF1 binds to specific Wnts and regulates the expression of cell cycle-related Wnt target genes for its growth-inhibitory effect on cancer cells. It was shown that WIF1 binds to Wnt1 and inhibits the growth of invasive bladder cancer cell lines via induction of G1 arrest. The G1 arrest by WIF1 is associated with down-regulation of SKP2 and c-myc and up-regulation of p21 and p27. In addition, SKP2, a substrate recognition component of Skp1-Cul1-F-box ubiquitin-ligase responsible for p27 protein abundance and a putative oncogene, is a potential target gene of the Wnt pathway.2 In addition, Wif1 plays important role in cardiomyogenesis. It was shown that the early upregulated Wnt inhibitory factor-1 (Wif1), stimulates cardiomyocyte differentiation in a similar manner as Wnt stimulation.4 WIF1 has also been implicated to play a role in normal prostate development. Furthermore, WIF1 was shown to be highly expressed in the developing and mature mouse skeleton and involved in osteoblast differentiation and chondrogenesis. Targeted deletion of mouse WIF1 augmented spontaneous and radiation-induced osteosarcoma formation.3 Moreover, Wif1 may play an essential role in the spatial and temporal regulation of Wnt signals in development of central nervous system.5
The commonly described effect of WIF1 on cancer cells is the inhibition of cancer cell growth. The Wnt/β-catenin pathway has been shown to determine the proliferation/differentiation balance through its regulation of G1-S transition in several cellular systems. The G1-S transition in cell cycle is driven mainly by cyclin-dependent kinase (CDK) 2 that is controlled by abundance of CDK inhibitors: p21/WAF1 (p21) and p27/Kip1 (p27). The regulation of G1-S transition is physiologically required for cell fate determination—a cell undergoing apoptosis, proliferation, or differentiation. However, during oncogenic transformation, G1-S transition is deregulated by enhanced oncogenic growth signaling and/or by loss of tumor suppressors, which then leads to overgrowth of transformed cells. Some Wnts (e.g., Wnt1) have shown oncogenic activities in both mouse models and cell cultures. The activation of the Wnt/β-catenin pathway by Wnts elicits specific target genes (e.g., c-myc and cyclin D1) for cell cycle regulation and growth. It is possible that the secreted Wnt antagonist WIF1 binds to specific Wnts and regulates the expression of cell cycle-related Wnt target genes for its growth-inhibitory effect on cancer cells. It was shown that WIF1 binds to Wnt1 and inhibits the growth of invasive bladder cancer cell lines via induction of G1 arrest. The G1 arrest by WIF1 is associated with down-regulation of SKP2 and c-myc and up-regulation of p21 and p27. In addition, SKP2, a substrate recognition component of Skp1-Cul1-F-box ubiquitin-ligase responsible for p27 protein abundance and a putative oncogene, is a potential target gene of the Wnt pathway.2 In addition, Wif1 plays important role in cardiomyogenesis. It was shown that the early upregulated Wnt inhibitory factor-1 (Wif1), stimulates cardiomyocyte differentiation in a similar manner as Wnt stimulation.4 WIF1 has also been implicated to play a role in normal prostate development. Furthermore, WIF1 was shown to be highly expressed in the developing and mature mouse skeleton and involved in osteoblast differentiation and chondrogenesis. Targeted deletion of mouse WIF1 augmented spontaneous and radiation-induced osteosarcoma formation.3 Moreover, Wif1 may play an essential role in the spatial and temporal regulation of Wnt signals in development of central nervous system.5
REFERENCES
1. Ai, L. et al: Carcinogenesis 27:1341-48, 2006
2. Tang, Y. et al: Mol. Cancer Therapeut. 8:458-68, 2009
3. Yee, D.S. et al: Mol. Cancer 9:162, 2010
4. Buermans, H.P.J. et al: PLos ONE 5:e15504, 2010
5. Hu, Y.A & Zhao, C.J: Zhejiang Da Xue Xue Bao Yi Xue Ban 39:93-6, 2010
2. Tang, Y. et al: Mol. Cancer Therapeut. 8:458-68, 2009
3. Yee, D.S. et al: Mol. Cancer 9:162, 2010
4. Buermans, H.P.J. et al: PLos ONE 5:e15504, 2010
5. Hu, Y.A & Zhao, C.J: Zhejiang Da Xue Xue Bao Yi Xue Ban 39:93-6, 2010
Products are for research use only. They are not intended for human, animal, or diagnostic applications.
Параметры
Cat.No.: | CP10277 |
Antigen: | Purified recombinant human WIF1 fragments expressed in E. coli. |
Isotype: | Mouse IgG1 |
Species & predicted species cross- reactivity ( ): | Human, Mouse, Rat |
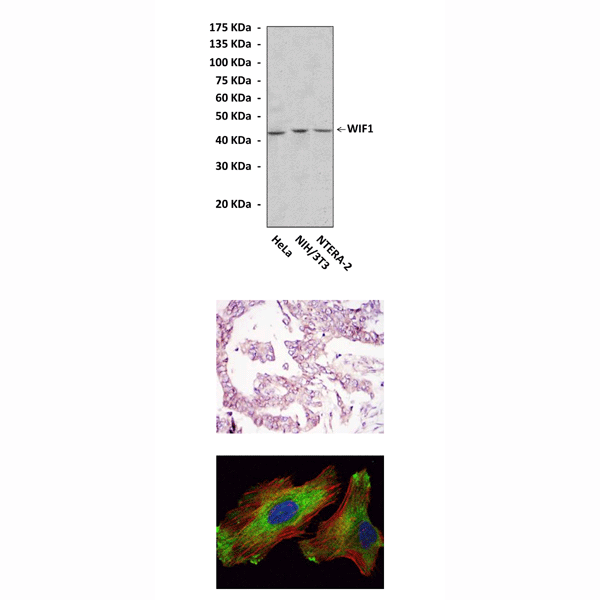
Applications & Suggested starting dilutions:* | WB 1:1000 IP 1:50 IHC 1:50 - 1:200 ICC 1:50 - 1:200 FACS n/d |
Predicted Molecular Weight of protein: | 43 kDa |
Specificity/Sensitivity: | Detects WIF1 proteins without cross-reactivity with other related proteins |
Storage: | Store at -20°C, 4°C for frequent use. Avoid repeated freeze-thaw cycles. |
*Optimal working dilutions must be determined by end user.
Документы

Информация представлена исключительно в ознакомительных целях и ни при каких условиях не является публичной офертой



