


Anti-RTN3: Mouse Monoclonal Reticulon 3 Antibody |
 |
BACKGROUND Reticulon (RTN) proteins are a group of membrane-bound proteins that are largely localized in the endoplasmic reticulum (ER), perhaps more restricted to the tubular ER. RTNs exist in plants, fungi, and animals. In mammalian genomes, four independent reticulon genes, rtn1 to rtn4, have been identified to encode a large number of gene products. All RTNs are composed of a variable length of N-terminal domain, which is completely divergent among RTN members, and a highly conserved RTN homology domain (RHD) that is unique to this family of proteins. Within the RHD, there exist two long stretches (28–36 amino acids) of hydrophobic residues, and each is expected to embed RTNs in the membrane. The divergent N-terminal domain among RTNs potentially allows each RTN member to exert its specific function. RTN4, or its more popular acronym Nogo, is identified as a critical inhibitory molecule in axonal growth and regeneration. Recently, reticulon proteins (including Nogo) have been shown to interact with beta-secretase, also known as beta-site APP-cleaving enzyme 1 (BACE1), and this interaction negatively modulates the enzymatic activity of BACE1. Among the RTNs, RTN3 has strong inhibitory effects on BACE1 activity and high expression by neurons. The first transmembrane stretch or domain (TM1) is important for the membrane insertion of RTN3. Disruption of the TM1 of RTN3 results in misfolding of the mutant protein and failure to integrate the protein into the membrane. Subtle changes in RTN3 membrane topology can disrupt its binding to BACE1 and its inhibitory effects on BACE1 activity.1 RTN3 is localized not only in the ER compartment but also in the Golgi, axons, dendrites, and growth cone. The localization of RTN3 in the neuritic region suggests its potential role in neuritic functions. It may be involved in membrane trafficking in the early secretory pathway. It has been demonstrated that RTN3 also plays a role in the formation of dystrophic neurites in Alzheimer brains. It was found that RTN3 is enriched in RTN3 immunoreactive dystrophic neurites (RIDNs) in AD brains. More importantly, transgenic mice overexpressing RTN3 (Tg-RTN3) develop RIDNs predominantly in their hippocampi, and this correlates with the formation of RTN3 aggregates in susceptible brain regions.2 Moreover, over-expression of RTN3 might increase DR5 surface protein and concomitantly more activate caspase pathways, which cause the c-FLIP cleavage and enhancement of TRAIL-mediated apoptosis.3 In addition, it has been shown that RTN3 participated in apoptosis regulation in other mechanisms. ER-bound RTN3 protein recruited endogenous FADD to the ER membrane and subsequently initiated caspase-8 cascade, including activation of caspase-8, processing of Bid and release of cytochrome c from mitochondria.4 CRELD1, a cell adhesion molecule played a critical role in atrioventricular septal defects, could partly change the localization of RTN3 from the endoplasmic reticulum to the plasma membrane and modulate the apoptotic activity of RTN3 through binding with it.5 RTN3 may also favor BCL2 translocation to the mitochondria upon endoplasmic reticulum stress.
REFERENCES
1. He, W. et al: J. Biol. Chem. 282:29144-51, 2007
2. Shi, Q. et al: J. Neurosci. 29:9163-73, 2009
3. Lee, J.T. et al: Cancer Lett. 279:185-92, 2009
4. Xiang, R. et al: Apoptosis 11:1923-32, 2006
5. Xiang, R et al: Mol. Cell. Biochem. 331:225-30, 2009
2. Shi, Q. et al: J. Neurosci. 29:9163-73, 2009
3. Lee, J.T. et al: Cancer Lett. 279:185-92, 2009
4. Xiang, R. et al: Apoptosis 11:1923-32, 2006
5. Xiang, R et al: Mol. Cell. Biochem. 331:225-30, 2009
Products are for research use only. They are not intended for human, animal, or diagnostic applications.
Параметры
Cat.No.: | CP10220 |
Antigen: | Purified recombinant human RTN3 fragments expressed in E. coli. |
Isotype: | Mouse IgG2a |
Species & predicted species cross- reactivity ( ): | Human |
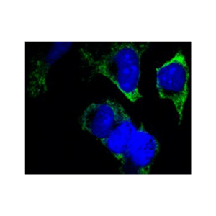
Applications & Suggested starting dilutions:* | WB 1:1000 IP n/d IHC 1:100 ICC 1:100 FACS n/d |
Predicted Molecular Weight of protein: | 113 kDa |
Specificity/Sensitivity: | Detects RTN3 proteins without cross-reactivity with other family members. |
Storage: | Store at -20°C, 4°C for frequent use. Avoid repeated freeze-thaw cycles. |
*Optimal working dilutions must be determined by end user.
Документы

Информация представлена исключительно в ознакомительных целях и ни при каких условиях не является публичной офертой



